澜海源创教育
标题:
为什么生物进化到今天是呼吸氧气而不是氮气?
[打印本页]
作者:
joe
时间:
2023-11-19 22:34
标题:
为什么生物进化到今天是呼吸氧气而不是氮气?
要回答这个问题,就要明白生物是怎么进化的?为什么好氧生物需要氧气?可以接收生物电子传递链上的电子的氧化性物质是怎么变化的,具有什么样的演变规律?生物与这些氧化性物质又是如何协同进化的?因为学过生物的都知道,所有细胞生物都有
电子传递链
,既然如此,那么所有的细胞生物就都需要氧化性物质来接收电子传递链上的电子。对上述问题的解答如下所述。
内容分以下几部分:1)为什么氧气可以作为生物的电子受体?2)生物进化模型;3)电子受体的演变特征;4)基于电子受体的生物进化模型
(一)为什么氧气可以作为生物的电子受体?
不难发现,所有细胞生物体都有一个共性,那就是都需要食物(或者成能量供体或
电子供体
),都需要氧化性物质用以接受电子传递链上的细胞代谢产生的电子。生物的代谢过程就是将食物中的电子传递给电子受体,即生物在自然界中就是一种电子传递
介体
或者说能量传递介体。个人认为,生物与其电子供体和电子受体相互影响,相互制约,对立统一,协同进化,任何一方发生变化,都会对“电子供体——生物体——电子受体”这个系统造成影响。
众所周知,氧气是几乎所有高等生物(如人类)唯一的电子受体。有意思的是,很多微生物,如地杆菌(
Geobacter
)和
希瓦氏菌
(
Shewanella
),可以选择多种氧化物作为电子受体,如 Fe(III)、Co(III)、U(VI)、Tc(VII)、
延胡索酸
、腐殖质等。不同的电子受体接受生物代谢产生的电子的方式往往不同。例如,当以水溶性的柠檬酸铁作为电子受体时(三价铁负责接受电子),有的
地杆菌
因细胞膜上缺少铁离子运输通道,致使铁离子主要在细胞壁和细胞膜之间的周质接受菌体代谢产生的电子;当以水难溶性的水铁矿(三价铁负责接受电子)作为电子受体时,由于水铁矿的分子量比较大而难以进入
细胞
,致使其主要在细胞外接受菌体代谢产生的电子。也有一些水溶性的氧化物(如延胡索酸等)或者
小分子氧化物
(如氧气),可以进入细胞内,因而主要在细胞内接受菌体代谢产生的电子。
一般情况下,电子受体越适合接受生物代谢产生的电子,就越容易驱动电子的流动,也就越有利于电子供体释放能量和传递电子,进而越适合环境向
能量均衡
状态转变的需求。此外,电子受体越有利于驱动电子在细胞内的流动,细胞中的“能量
货币
(ATP 等)”就越充足,细胞也就越有充足的能量来驱动自身能量传递能力的提升。总之,对于生物而言,电子受体越合适,就越有利于生物能量传递能力的发挥,也就越有利于生物自然价值的呈现。对于电子受体而言,越容易接受生物代谢产生的电子,以其作为电子受体的生物的数量和种类就会越多,进而表现为各生物之间对电子受体的竞争越激烈。
水难溶性的电子受体主要存在于
地球
表面的土壤或淤泥中。这类电子受体的空间流动性(扩散能力)相对比较差,但在一个特定环境中的位置却相对比较稳定。水溶性的电子受体或分子量较小的电子受体主要分布在水环境或大气环境中。这类电子受体的空间流动性相对比较强,这使得其在一个特定环境中的分布相对更容易实现均衡。以难溶于水的铁矿石和分子量相对较小的氧气为例,铁矿石在地球表面的分布是不均匀的,而且自然因素(如风)难以影响其在地球表面的分布,这使得其在特地区域的分布又是相对稳定的。相比较而言,
氧气分子
的流动性就比较强,而且自然因素(如风)很容易带动氧气分子的流动,不过由于氧气分子的分子量相对比较小,氧气分子的
布朗运动
又使得其在空气中的分布很容易的再次实现均衡。
细胞复杂的结构组成说明,生物的起源过程需要在一个相对稳定的环境中发生。环境越稳定,组成生物的各种组分之间的联系就越不容易被破坏,也就越有利于生物的形成。由此可进一步推测,生物的起源过程及形成初期,所在特定环境内的电子受体也应该是稳定存在的。
难溶性电子受体
在特定环境中分布的均衡性和稳定性,使得其更利于非生物系演变成生物。相比较而言,空间流动性更好的电子受体,因其在大空间尺度上分布更加均匀且更容易利用,因而更利于生物的增殖和传播。
可能有人会问,为什么现在大多数的生物,特别是陆生生物,会选择氧气作为电子受体,而不是其它氧化物?为什么这些生物只能以氧气作为唯一的电子受体?既然这是已经发生的事实,推测这一事实出现的原因主要有以下几点:
1)氧气可以通过自由扩散的方式进入细胞,使得其可以相对更容易的接受生物代谢产生的电子;
2)氧气接受电子的能力相对于其它氧化物而言更强,同时,对细胞造成的氧化毒害又相对较小;
3)“自养型”生物在二氧化碳等物质的固定过程中,实现了
氧元素
以分子形式从化合物中的分离(如植物的光合作用等),导致自然界中氧气含量的增加;
4)自然因素(如火山、雷电等)致使氧元素以分子形式从其化合物中分离,同样导致自然界中氧气含量的增加;
5)氧元素以分子的形式从化合物中脱离出来,也是降低其原来所在化合物能量的过程,符合自然界在能量流动方面的需求;
6)氧元素是地球表面含量最多的元素之一,为氧气的大量出现及更多的生物选择氧气作为电子受体提供了可能。
总之,氧气含量高、分布广、流动性好及其易于接受电子的属性,是越来越多的生物选择以其作为电子受体的根本原因,这同样也为生物的进化提供了驱动力。
(二)生物进化模型
能流进化论,或称能流矛盾论的意义在于,通过解析生物的能量供体(食物)、能量受体(环境中用以接收流经电子传递链的细胞代谢所产生的电子的氧化性物质,如氧气)以及二者与生物组成的能量传递单元在达到能量流动平衡时的条件,就可以通过监测环境中能量供体和能量受体的变化,揭示或预测生物在环境中的演变方向、演变方式、演变缘由及与其它种类生物之间的关系。此外,通过解析生物调整其能量传递能力的方式,也可以预测该物种随环境中能量供体和能量受体丰度的变化而呈现的变化趋势。
基于能量供体、能量受体及生物的能量传递能力在历史长河中的演变,以及三者之间的矛盾关系,可以构建一个三维坐标系(X 轴:能量供体;Y 轴:能量受体;Z 轴:物种的能量传递能力)。通过这个坐标系,也可以预测或揭示生物的进化方向及物种之间的相互关系。在这个坐标系中,两个物种的位置越近,说明它们对应的能量供体和能量受体共享率越高,那么,它们之间对能量供体和能量受体的竞争也就越强烈。如
图 1
所示,物种 1 和物种 2 之间对能量供体和能量受体的竞争要比它们分别与物种 3 之间的竞争更加激烈。
图 1 基于能量供体、能量受体和生物能量传递能力的演变方向和能流矛盾论构建的坐标系
(三)电子受体的演变特征
如果一个环境不够稳定,生物各个部分之间的相互联系和相互作用就难以维持,也就不可能有生物的出现。水难溶性电子受体的空间稳定性使得组成生物的各个组分或系统之间更容易相互合作,生物的能量传递能力也能得以稳定的发挥。由此可见,稳定的电子受体环境是生物起源的必要条件,本书认为这一环境便是富含水难溶性电子受体的环境。
然而,由于水难溶性电子受体的空间流动性比较差,随着生物的增殖,生物之间对电子受体的竞争必然越来越激烈。此外,随着电子的不断接受,特定环境中的电子受体对电子的接受能力越来越弱。空间流动性差这一特点,致使电子受体难以在该特定区域得到有效补充,这将进一步加剧生物之间对电子受体的竞争。在这个过程中,特别是当电子受体的电子容纳量快达到饱和时,如果环境中的电子供体含量充足,那么生物就不得不选择其它可以利用的氧化性物质作为电子受体。
在自然因素(如火山喷发、雷电等)及生物因素(如生物的代谢产物、降解产物等)的影响下,水溶性氧化物的数量和种类逐渐增多。这类氧化物的空间流动性相对比较好,因而不适合非生物体系向生物的演变。然而,流动性好这一特点,使得可溶性氧化物得以在更大的空间尺度上分布,进而使得生物在特定空间里对电子受体的竞争相对缓和。这是因为,以水难溶性氧化物作为电子受体的生物,其只能分布在这些氧化物的表面,而对于具有相同电子接受能力的水溶性氧化物而言,电子受体的分布范围则是整片水域,因而生物的分布范围在理论上也是整片水域。就像一个开口的装满白砂糖的瓶子,假设瓶口的位置最多只能供 100 只蚂蚁取食,如果将瓶子里的白砂糖全部倒出来,并平摊在一张比瓶口面积大 100 倍的桌面上,对于相同数量的蚂蚁而言,其取食空间也就增大了 100 倍,也就意味着可以有更多的蚂蚁同时享用白砂糖。
再者,有的水溶性氧化物,特别是能进入细胞内的氧化物,相对于水难溶性的氧化物而言,更容易接受生物代谢产生的电子,这就使得一部分生长在水难溶性电子受体表面上的生物逐渐适应并能够利用这类水溶性氧化物作为电子受体。水溶性电子受体提高了生物代谢产生的电子在生物中的传递效率,进而使得更多的能量被生物释放和传递,这适合自然界对生物所属能量传递单元的能量平衡能力的需求,相应的生物的能量传递能力也就有存在的可能。此外,经生物释放和传递的能量越多,生物可获取的用以提升自身能量传递能力的能量也就越多。这些能量不仅用于生物自身结构组成的升级优化,还可用于提高增殖速度。生物数量的增加,表现为单位空间内生物对电子供体和电子受体的竞争加剧。在竞争压力下,有些生物通过对自身结构的升级,如产生鞭毛等运动器官,提升了自身的空间移动能力,即运动性,因而可以主动的移动至竞争环境相对缓和的空间。此外,也有些竞争能力相对较强的生物,可以利用自身的攻击系统去消灭或抑制竞争对手,以获取自身相对更大的生存空间。总之,水溶性氧化物作为电子受体,驱动了生物从地表或地下环境进入湖泊或海洋。在这个过程中,生物的增殖能力和扩散能力也都得到提升。此外,水溶性电子受体的多样性和更广的空间分布性还推动了生物在结构和功能方面的多样化升级,最终表现为物种的多样化。
水溶性电子受体的多样性和更广的空间分布性推动了生物在结构和功能方面的多样化升级,最终表现为物种的多样化。在新出现的物种中,有些还保留着自养系统,只是因为周边环境的不合适,致使其自养系统一直处于休眠状态。当这类生物重新处于适合其自养系统正常运行的环境中时,自养系统有可能会被重新激活。例如,一些“光能自养型”生物因环境变化而进入了没有光的环境,在对新环境的适应过程中,一直保留着原来的光能自养系统,只是为了节约能量而将其光能自养系统处于休眠状态。在与电子受体的共进化过程中,有的生物重新回到了可以接受光能的环境中,如水面。充足的光照使得这些生物中的光能自养系统再次被激活,甚至有可能得到进一步的完善和升级。以光能作为能量供体的生物的种类和数量的增加,加剧了光能自养型生物之间对光能的竞争。有的生物可能通过提高光能利用率来减缓自身在光能获取方面的竞争弱势,有的生物则可能通过增强自身组织细胞的增殖来增加光接收面积,进而提升自身在光能获取方面的竞争优势。
除了自养系统,生物内也有可能存在一些对光照敏感的组分或系统。这类系统在受到光照后,有的功能可能会受到影响,有的则可能会呈现出新的功能。对于功能受到影响的系统,生物为了维持其正常的能量传递能力,就需要对其防御系统进行改造升级,以尽可能的降低或摒除光照对这些光敏感系统的影响。例如,有些化合物在光照条件下会发生共价键的均裂(共用电子均等地分配给成键的两个原子的过程),均裂后的两个部分各带有一个未成对电子,是一种活泼中间体。这类中间体又被称为自由基或游离基。如果自由基的含量比较高,就会对生物的健康造成影响。自由基的种类很多,比较常见有氢自由基、氯自由基、甲基自由基、氧自由基等。以超氧自由基为例,生物为了尽可能的降低超氧自由基对自身系统的伤害,会产生一种叫做超氧化物歧化酶的蛋白,该蛋白可将有害的超氧自由基转化为过氧化氢。过氧化氢在过氧化氢酶的催化作用下立即分解为无害的水。
对于激发出新功能的系统,如果该功能有利于提高生物的能量传递能力,将有可能会被生物加以优化利用。以视觉系统为例,视觉系统可以让生物更容易的探寻到能量供体或能量受体,以及躲避来自竞争对手的伤害等。鉴于视觉系统的这些优点,很多物种在进化过程中也在不断的对自身的视觉系统进行升级改造,使其更好的为自己服务。例如,乌贼的眼睛能够很好地处理海洋中浑浊的光线,使其在海洋中可以看得更清楚;比目鱼的两只眼睛位于同一侧,使其在侧躺在海床上时,更容易发现和捕捉到猎物,同时又可避免沙子进入眼睛;蜻蜓和苍蝇的眼睛是由数万只单眼组成的复眼,这种眼睛可以有效的计算自身与所观察物体的方位,从而有利于作出更快速的判断和反应。除了对与这类新功能直接相关的系统进行结构和功能的升级外,有的生物为了让这个新功能得以更充分的施展,还会驱动一些相关系统结构和功能的升级。例如,很多植物能够通过调节生长激素在自身不同组织器官的分布量,来改变组织器官相对太阳的朝向,进而使得自身可以获取更多的光照,该进化结果便是植物等生物所呈现出来的趋光性。
在水溶性氧化物可以作为生物电子受体的初期,水溶性氧化物的种类和数量繁多,但却还未出现一种占有主导地位的电子受体。此时,充足的电子供体和种类繁多的电子受体驱动了生物种类的爆发。这可能是寒武纪生物大爆发的原因之一。
在这些可被生物用作电子受体的水溶性氧化物中,有的丰度越来越高(如铁离子),同时,以其作为电子受体的生物的种类和数量也越来越多,该电子受体参与的能量传递单元的能量平衡能力也越来越强。这类电子受体逐渐成为生物的主导电子受体。与此同时,以主导电子受体作为电子受体的生物,也逐渐成为一个特定环境中共享“电子供体”的物种中的优势物种。在优势物种与劣势物种对“共享电子供体”的竞争中,劣势物种的劣势越来越明显,表现为能量传递能力越来越弱。共享“电子供体”的物种中的劣势物种,如果不能适应并利用其它物质作为能量供体,那么,随着时间的推移,该物种可能会被自然界所淘汰。反之,如果该劣势物种可以利用其它种类的能量供体,根据自然界会以当前最高效的方式使自身的能量向均衡状态转变可推知,这种能够适应其它电子供体的劣势物种有很高的几率会存活下来。随着利用主导电子受体的生物的数量和种类的增加,共享主导电子受体的物种之间对主导电子受体的竞争也日益激烈。
水溶性电子受体驱动着生物在地球表面更广阔的范围内分布和传播。在这个过程中,光能自养型生物的增殖和传播,使得空气中氧气的含量不断地增加。由于氧气能够通过自由扩散的形式进入细胞内,且接受电子的能力相对更强等优势,越来越多的生物能够适应并逐渐选择氧气作为电子受体。然而,氧气难溶于水的特性,致使其在空气中的含量比在水中的含量更高。总之,氧气作为生物的电子受体后,一方面使得生物的能量传递能力得到进一步提升,一方面其水难溶性也驱动着生物逐渐从海洋走向陆地。
(四)基于电子受体的生物进化模型
基于电子受体的特征及其与生物之间关系的讨论,《隐藏的动力:生物在自然界中的价值》一书构建了一种电子受体驱动的生物进化模型(
图 2
),即电子受体在生物进化过程中的演变历程被认为是:首先,难溶于水的、在特定区域内稳定存在的、相对难以被利用的氧化物(如水铁矿、磁铁矿等)是生物最初阶段的主要电子受体;随着水溶性氧化物的含量及种类越来越多,因其相对更容易接受生物代谢产生的电子,且生物对其竞争相对缓和等原因,越来越多的生物逐渐适应并选择这类氧化物作为电子受体;随着氧气的大量产生,因其氧化性更强、分子量更小、分布更均匀、更容易接受生物代谢产生的电子等特性,致使越来越多的生物逐渐适应并选择氧气作为电子受体。
图 2 驱动生物进化的电子受体的演变
该进化模型的构建之所以选择电子受体作为参照而非电子供体,主要是基于以下两件事实:
1)对于任一生物而言,电子供体(食物)的多样性往往多于电子受体的多样性。例如,人可以吃很多种食物,如肉类、蔬菜、水果等,但只能以氧气作为电子受体。当然,自然界中也存在着一些专食性的物种,如大熊猫只吃竹子、考拉只食用桉树叶子、蜗鸢专吃蜗牛等。然而,对于专食者而言,其专食性也是相对的,在偏好的食物匮乏时,它们也会选择其它食物,例如,野生的大熊猫也会吃草、野果、昆虫、竹鼠、牛羚、甚至附近村落里养的羊和垃圾堆的剩饭菜等。
2)电子受体的缺乏对生物造成的影响一般要比电子供体缺乏造成的影响更严重。例如,人可以三天不吃东西,但不能 1 小时不呼吸;冬眠中的青蛙可以几个月不吃不喝,但冬眠的环境中不能没有氧气等。
(五)能流进化论的特色
最近在利用能流进化论解释各种生物现象及生物行为时,也在不断总结和思考能流进化论的特色和优势是什么?与其它进化论的区别主要体现在什么地方?在此,特将近期的思考整理一下。
1.能流进化论是从自然界能量流的角度探究生命的起源与进化问题,而其它进化论主要从构成细胞的物质的角度探究生命的起源与进化。生态系统中有循环的物质流、双向的信息流和单向的能量流三种主要的功能流,从中也可以发现,能量流相对物质流和信息流更简单。天下难事,必作于易(道德经第六十三章),因而从能量流的角度探究生命的起源与进化问题是值得推崇和应用的方法,也是相对更为合理的方法。
2.能流进化论是从生物在自然界中价值的角度探究生命的起源与进化问题,而其它进化论主要是从组成生物体的物质分子在生物体中的价值的角度探究生命的起源与进化问题,因而才有了生命是起源于 DNA、RNA 还是蛋白质等争议话题。不识庐山真面目,
只缘身在此山中
(苏轼,题西林壁)。从自然界的角度探究生命的起源与进化问题比从生物体的角度要相对更系统、更全面、更合理、更准确。
3.能流进化论的论述方法是从自然界中的能量流聚焦到有能量传递介体介导的能量流,根据所有生物体都有介导能量传递这一共性及自然界对能量均衡分布的需求,推导非生命态能量传递介体向生命态能量传递介体演变的原因、过程和动力,该过程即为生命的起源。基于生命起源,继续根据能量传递介体在自然界中的价值共性,推究生物的进化。由此可见,在能流进化论中,生物进化是基于生命起源的生物进化,生命起源又是基于自然界能量流的生命起源,三者因能量流密切衔接,层次分明。然而,目前的很多进化论主要关注物种的起源及生物的进化,逃避生命起源问题,给人一种“头重脚轻根底浅,嘴尖皮厚腹中空”的感觉。
4.能流进化论认为,能量供体(食物)、生物体和能量受体(接收流经电子传递链的细胞代谢产生的电子的氧化性物质)三者之间相互影响,相互制约,对立统一,协同进化。从能流矛盾的角度,可以解释自然选择的依据是什么?适者生存中的“适者”是指什么等问题。然而,其它进化论则主要从生物的角度探究生物的进化。
5.能流进化论的目的不只是阐述生命的起源与进化问题,更在于其应用性,没有应用价值的理论是没有灵魂的理论。能流进化论可以用于解释各种生物现象及生物行为,也还可以用于审视目前存在的认识、假说等。例如在前述博文中,利用能流进化论解释了恐龙灭绝的原因、寒武纪物种大爆发的原因、爱尔兰大饥荒的原因、癌症的发生和转移原因、心脏很少癌变的原因、果树修剪的原因、下雨前燕子低飞和蚂蚁搬家的原因等,解释了医学的进步与自然选择是否冲突的问题、外星生命问题等,还利用能流进化论提出了电子传递链的新的运行机制……这些是其它进化论所欠缺的。
能流进化论是《
隐藏的动力:生物在自然中的价值
》(汕头:
汕头大学出版社
,2020-12,CIP2020268179,ISBN 978-7-5658-4304-4.)一书阐述的一种分析问题的方法论,也是一本基于
热力学定律
,从生物在自然界能量流动方面价值的角度阐述生命起源与进化问题的书。该书共分为六章,第一章介绍了从能量和价值的角度探讨生命起源与进化问题的缘由;第二章介绍了自然界中能量流动所遵循的基本定律及生物在自然界能量流动中的价值;第三章从生物在自然界能量流动中价值的角度,探讨了生物是如何从
非生命态
一步步演变为生命态的;第四章从生物在自然界能量流动中价值的角度,探讨了生物的演变缘由、演变方式和演变方向;第五章提出了“能流进化论”学说,借以阐释了“物竞天择,适者生存”现象;最后一章通过实例探讨了
能流矛盾论
在农业、医学、经济等领域的应用。
《
隐藏的动力:生物在自然界中的价值
》(汕头:汕头大学出版社,2020-12,CIP2020268179,ISBN 978-7-5658-4304-4.)一书电子稿可从该链接
https://blog.sciencenet.cn/blog-3479210-1308965.html
免费下载。
欢迎光临 澜海源创教育 (https://lhycedu.com/)
Powered by Discuz! X3.5
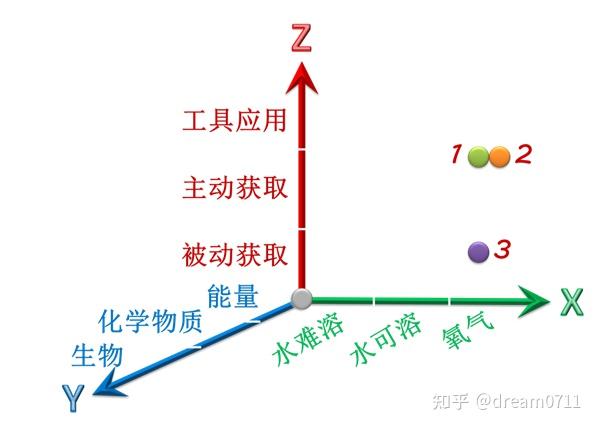 图 1 基于能量供体、能量受体和生物能量传递能力的演变方向和能流矛盾论构建的坐标系
图 1 基于能量供体、能量受体和生物能量传递能力的演变方向和能流矛盾论构建的坐标系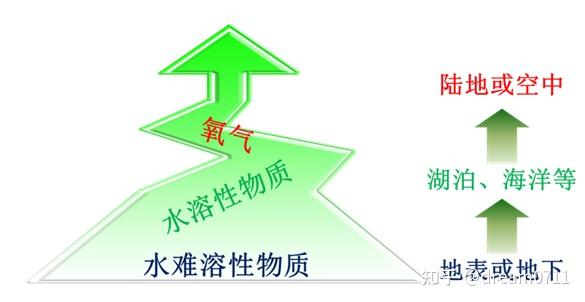 图 2 驱动生物进化的电子受体的演变
图 2 驱动生物进化的电子受体的演变